
Introducción
La industria avícola global se enfrenta a desafíos económicos de gran magnitud derivados de la contaminación de materias primas y piensos por micotoxinas (Chowdhury et al., 2005; Girish and Smith, 2008). Dentro del sector aviar, el pavo (Meleagris gallopavo) destaca por ser una de las especies con mayor susceptibilidad a estos compuestos, con ejemplos de especial interés como la aflatoxina B1 (AFB1).
Aflatoxinas
La identificación inicial de las aflatoxinas estuvo intrínsecamente ligada a la patología aviar. El brote masivo de la denominada «Enfermedad X del Pavo» ocurrido en la década de los 60 en el Reino Unido, con una mortalidad superior a los 100.000 ejemplares de pavo, definió un cuadro clínico de anorexia, letargo y cuadros severos de necrosis y hemorragia hepática. La investigación posterior confirmó a Aspergillus flavus como el agente contaminante del pienso, permitiendo a partir de este momento la caracterización de las aflatoxinas B1, B2, G1 y G2.
La excepcional sensibilidad del pavo doméstico a la AFB1 radica en una particularidad genética que limita su capacidad de detoxificación: la capacidad limitada de las enzimas glutatión S-transferasas (GST) para neutralizar los metabolitos reactivos de la toxina. Tras la ingesta, la AFB1 experimenta un proceso de bioactivación hepática que genera el metabolito exo-AFB1-8,9-epóxido (AFBO). Este metabolito electrofílico posee un alto potencial carcinogénico y mutagénico, siendo incluso superior al de la AFB1. Debido a la ineficiencia de las enzimas GST hepáticas, el AFBO no se neutraliza eficazmente, derivando en un daño celular severo, una marcada inmunosupresión y un compromiso sistémico de la salud del ave (Kim et al., 2013; Reed et al., 2018; Jorud et al., 2024).
Clínicamente, la exposición a aflatoxinas se traduce en un deterioro inmediato de los índices productivos, con caídas drásticas en la tasa de crecimiento, en la eficiencia alimentaria y en la producción de huevos (Aviagen Turkeys, 2018; Monson et al., 2014; Tilley et al., 2017; Nava-Ramírez et al., 2024). El hígado, principal órgano diana, manifiesta alteraciones macroscópicas que incluyen esteatosis, fibrosis y una coloración pálida o amarillenta característica (Monson et al., 2014; Aviagen Turkeys et al., 2018; Nava-Ramírez et al., 2024). Más allá de la hepatotoxicidad, la AFB1 actúa como un potente inmunosupresor que incrementa la susceptibilidad a patologías secundarias y compromete la integridad de la barrera intestinal, induciendo disbiosis y un aumento de la permeabilidad de la mucosa (Monson et al., 2014; Maguey-González et al., 2024; Reed et al., 2018). También se ha observado que en pavos las aflatoxinas interfieren en el metabolismo de la vitamina D, lo que reduce la fuerza de los huesos y provoca debilidad en las patas (Aviagen Turkeys et al., 2018).
A nivel molecular, la toxina interfiere en la expresión de genes críticos vinculados a la apoptosis y al metabolismo lipídico. Estos efectos sistémicos pueden iniciarse incluso durante el desarrollo embrionario y manifestarse mediante alteraciones en la bioquímica sanguínea o variaciones en el peso relativo de órganos linfoides esenciales, tales como el bazo y la bolsa de Fabricio (Monson et al., 2016; Tilley et al., 2017; Nava-Ramírez et al., 2024).
Tricotecenos: deoxinivalenol y toxina t-2
Los tricotecenos, como el deoxinivalenol (DON) y la toxina T-2, centran su patogenicidad en la disrupción de tejidos con una elevada tasa de proliferación celular y recambio proteico. El mecanismo de acción molecular fundamental de ambos metabolitos radica en la inhibición de la síntesis proteínas, ARN y ADN mediante su unión específica a la subunidad ribosomal. Este bloqueo de la traducción proteica impide la división celular y consolida a los tricotecenos como contaminantes altamente disruptivos para la fisiología y el rendimiento productivo de los pavos (Chowdhury et al., 2005; Girish and Smith, 2008; Hassan et al., 2011).
En el tracto digestivo, ambas toxinas alteran drásticamente la arquitectura de la mucosa, provocando vellosidades más cortas y delgadas en el duodeno y el yeyuno, lo que reduce significativamente el área de absorción efectiva de nutrientes y la eficiencia alimentaria (Girish and Smith, 2008).
Específicamente, el DON, también llamado vomitoxina, compromete la función digestiva al inhibir los cotransportadores de Na+-D-glucosa y Na+-L-prolina, lo que deriva en una merma de la ganancia media diaria de peso (GMD), especialmente durante las fases de iniciación. A nivel sistémico, actúa como un potente agente inmunomodulador que eleva los niveles de IgA biliar hasta 4,45 veces, evidenciando un estrés inmunológico activo en las mucosas que incrementa la susceptibilidad ante agentes infecciosos. Es necesario subrayar que la toxicidad del DON en pavos en condiciones de campo suele verse aumentada por su co-ocurrencia sinérgica con otras micotoxinas, como la zearalenona (ZEA) y el ácido fusárico (Hassan et al., 2011).
Por otro lado, la toxina T-2 destaca por su extrema citotoxicidad y su carácter irritante. En cuadros de toxicosis aguda, se asocia con el síndrome de muerte súbita, mostrando necropsia, hemorragias generalizadas y un enrojecimiento en los músculos de la pechuga (Hassan et al., 2011). También se puede observar la aparición de lesiones orales (Figura 1) y erosiones en la molleja (Aviagen Turkeys et al., 2018). A nivel histológico, la toxina T-2 actúa como un potente inmunotóxico que induce procesos de apoptosis y necrosis en órganos linfoides primarios como el timo y el bazo, comprometiendo de forma severa la competencia inmunitaria del pavo (Girish and Smith, 2008).
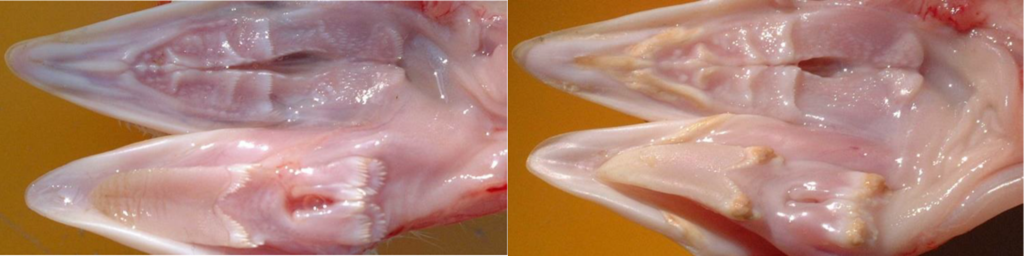
Figura 1. Lesiones orales y necrosis causadas por la toxina T-2. A la derecha cavidad oral normal, a la izquierda cavidad oral afectada (Aviagen Turkeys et al., 2018).
Fumonisinas
Al igual que en el resto de especies avícolas, en los pavos las fumonisinas actúan principalmente a través de la interrupción de la biosíntesis de los esfingolípidos mediante la inhibición competitiva de las enzimas ceramida sintasas (CerS) (Guerre et al., 2022). Esta interferencia bioquímica bloquea la conversión de esfinganina en ceramidas, provocando una acumulación de esfinganina libre que eleva drásticamente el ratio esfinganina:esfingosina (Sa:So) en el hígado y el plasma. Este parámetro se consolida como el biomarcador diagnóstico de referencia para monitorizar la exposición a estas toxinas en condiciones de campo (Tardieu et al., 2019).
En el pavo y otras aves de corral, la respuesta molecular frente a las fumonisinas presenta una dinámica distintiva: se observa una reducción de las ceramidas de cadena media en contraste con un incremento de las de cadena muy larga y de los niveles de dihidroceramidas. Esta configuración metabólica específica, sumada a la ausencia de activación de las vías de reciclaje de la esfingomielina, explicaría la mayor resistencia que muestran estas especies frente a las fumonisinas en comparación con el ganado porcino (Guerre et al., 2022).
Sin embargo, la exposición crónica no está exenta de consecuencias patológicas severas, manifestándose en retrasos del crecimiento, cuadros de inmunosupresión y edema pulmonar. En episodios de toxicidad aguda, las lesiones observadas incluyen esplenomegalia, hepatomegalia y hemorragias en el parénquima renal (Hassan et al., 2011). Es fundamental advertir que el riesgo sistémico se multiplica ante la co-ocurrencia sinérgica con la AFB1 o la toxina T-2, interacciones que alteran el perfil bioquímico plasmático y elevan la tasa de mortalidad (Tilley et al., 2017).
Desde una perspectiva de salud intestinal, las fumonisinas comprometen la integridad del epitelio, facilitando la translocación de patógenos oportunistas. Un factor crítico a considerar en la gestión de riesgos es la farmacocinética de la fumonisina B1 en pavos. A pesar de una eliminación plasmática rápida, muestra una persistencia prolongada en el tejido hepático, con una vida media de eliminación estimada en 124 horas. Esta bioacumulación en hígado y tejido muscular subraya la importancia de implementar estrategias de control eficaces para garantizar la seguridad alimentaria y la sanidad animal (Tardieu et al., 2019).
Ocratoxina A
La ocratoxina A (OTA) se posiciona como una de las micotoxinas de mayor impacto crítico para la industria avícola, actuando como un potente agente nefrotóxico y hepatotóxico en pavos. Su mecanismo de acción citotóxico se fundamenta en la inhibición de la síntesis de proteínas, ADN y ARN, proceso que se ve agravado por la estimulación de la peroxidación lipídica a nivel celular. Debido a su elevada afinidad por las proteínas séricas, la OTA presenta una farmacocinética de distribución persistente, acumulándose en concentraciones biológicamente activas en riñones, hígado, sangre y en el contenido del yeyuno. En pavos jóvenes, incluso niveles mínimos en la dieta pueden comprometer la integridad orgánica y erosionar drásticamente la eficiencia productiva (Mazur-Kuśnirek et al., 2024).
Fisiopatológicamente, el cuadro de intoxicación por OTA se manifiesta mediante una marcada hepatomegalia y nefromegalia, acompañadas de hemorragias generalizadas que afectan también al bazo y al parénquima pulmonar. Este deterioro sistémico tiene un reflejo directo en la productividad, con reducciones significativas en el peso corporal final y en el consumo voluntario de pienso. Asimismo, la exposición crónica altera el perfil bioquímico sanguíneo, induciendo una hipoproteinemia y una disminución de las gamma-globulinas séricas, indicadores claros de un compromiso severo del metabolismo proteico y de la capacidad de síntesis hepática. A nivel digestivo, se han documentado alteraciones en el pH en el proceso de digestión en secciones críticas como el ciego, lo que sugiere una disrupción del equilibrio luminal (Hassan et al., 2011; Mazur-Kuśnirek et al., 2024).
El impacto de la OTA sobre el sistema inmunitario es determinante para la rentabilidad de la explotación. Como agente inmunotóxico, deprime la inmunidad celular y humoral, reduciendo la actividad de efectores clave de la inmunidad innata, como la lisozima y la ceruloplasmina séricas. Esta brecha inmunitaria no solo incrementa la susceptibilidad de los pavos frente a patógenos oportunistas, sino que compromete la respuesta a los programas de vacunación, invalidando las estrategias de medicina preventiva en la granja (Chowdhury et al., 2005; Mazur-Kuśnirek et al., 2024).
Zearalenona
La zearalenona (ZEA) se define como una micotoxina de tipo estrogénico. En condiciones de campo, su detección es recurrente como co-contaminante en cereales, coexistiendo habitualmente con el DON, lo que complica el diagnóstico clínico debido a las interacciones entre metabolitos (Chowdhury et al., 2005). En comparación con las especies porcinas, los pavos manifiestan una notable resiliencia frente a esta toxina en términos de parámetros zootécnicos. Investigaciones con dosis experimentales elevadas han confirmado que la ZEA no compromete de forma significativa la ganancia media diaria de peso, el índice de conversión ni el peso relativo de órganos diana, incluidos los testículos y los ovarios (Tilley et al., 2017).
A nivel metabólico, la exposición a niveles convencionales de ZEA no induce alteraciones en el perfil de esfingolípidos hepáticos. Su impacto fisiológico suele quedar subestimado o enmascarado por la toxicidad más agresiva de otras micotoxinas presentes en las mezclas naturales del pienso (Guerre et al., 2022). No obstante, la ingesta de concentraciones masivas puede desencadenar una respuesta estrogénica y alteraciones etológicas específicas. En machos expuestos a niveles críticos, se ha documentado un comportamiento de «pavoneo» (strutting) exacerbado, vinculado a un incremento en la turgencia y coloración de las carúnculas, así como a procesos inflamatorios en la cloaca (Tilley et al., 2017).
Desde la perspectiva inmunológica, aunque la ZEA de forma aislada presenta un impacto limitado, su inclusión en dietas naturalmente contaminadas puede favorecer una depresión moderada de la inmunidad mediada por células, afectando específicamente la respuesta de los linfocitos T CD8+ (Chowdhury et al., 2005; Tilley et al., 2017).
Conclusiones
Los piensos contaminados por micotoxinas son una amenaza para la producción de pavos, ya que estas provocan graves problemas de salud que afectan negativamente al rendimiento y al bienestar de las aves. Debido a su impacto es fundamental aplicar protocolos de control estrictos y medidas eficaces que logren reducir los daños y proteger la rentabilidad de la granja.
A pesar de que los efectos de las micotoxinas son conocidos, todavía es necesario profundizar en la investigación específica para pavos. Necesitamos entender mejor cómo funcionan sus mecanismos de toxicidad y definir umbrales de seguridad más precisos. Solo con un conocimiento técnico sólido podremos desarrollar estrategias de manejo y soluciones de prevención que respondan realmente a las necesidades de esta especie y aseguren un estatus sanitario óptimo.


