
Mycotoxin-Induced Alterations in Motility, Rumination, and Neuromuscular Function
Impact of mycotoxins on the gastric compartments of the ruminant
Introduction
The reticulum is the smallest and most cranial compartment of the ruminant gastric system, with a capacity of only 7–12 liters in adult cattle. However, its modest volume is in stark contrast with its functional relevance within digestive physiology: without the reticulum, there is no rumination. Without rumination, there is no salivary buffer. Without a salivary buffer, the rumen acidifies, the microbiome collapses and digestive efficiency plummets. This functional dependency makes the reticulum the initial link in a chain of events that can culminate in subacute ruminal acidosis (SARA), severe dysbiosis and systemic immune compromise (Mattioli, 2003; Pittroff & Kothmann, 2025).
Unlike the rumen, which has received the greatest research attention regarding mycotoxins in ruminants, the reticulum has received little attention as a specific target for these toxins. This neglect has practical consequences: the effects of mycotoxins on reticular motility and the rumination reflex are silent, subclinical and difficult to quantify under field conditions, but their repercussions on productive efficiency and health are documentable and significant. This article reviews the anatomy, histology and physiology of the reticulum from the perspective of mycotoxin-induced damage, with emphasis on the neurotoxic, inflammatory, and motor mechanisms that explain the observed dysfunction.
The most relevant mycotoxins affecting the reticulum include T-2 toxin and other type A trichothecenes (characterized by high cytotoxicity and neurotoxic activity), aflatoxin B1 (AFB1) (with neuro-hepatic metabolism), zearalenone (ZEN) (with endocrine-motor effects) and deoxynivalenol (DON) (type B trichothecene). The latter, in addition to its digestive impact, has neurotoxic capacity by altering neurotransmitter levels and crossing the blood-brain barrier, directly affecting motility and appetite (Zhang et al., 2020). Exposure to these toxins, a common situation in practical feeding, generates synergistic effects that amplify the individual damage of each compound (Fink-Gremmels, 2008; Mostrom & Jacobsen, 2020).
1. Anatomy and Histology of the Reticulum
The reticulum is located immediately caudal to the diaphragm, in close contact with the caudal surface of the diaphragm and the liver. Its proximity to the pericardium explains traumatic reticular pericarditis, a complication of reticulitis. The reticulum is separated from the rumen by the rumino-reticular fold as seen in Figure 1, permeable to fluid and fine particles, forming together with the rumen a morphofunctional unit: the reticulo-rumen (Mattioli, 2003; Febres & Vergara-López, 2007).
The most distinctive histological feature of the reticulum is its mucosa organized in primary, secondary and tertiary muscular folds that intersect forming polygonal cells resembling the alveoli of a honeycomb. The ridges of the folds are covered by keratinized conical papillae. This honeycomb architecture increases the contact surface and gives the reticulum its unmistakable appearance. The musculature of the reticulum is well developed, with layers of smooth muscle oriented in different planes that allow it to perform powerful, multidirectional contractions (Mattioli, 2003).
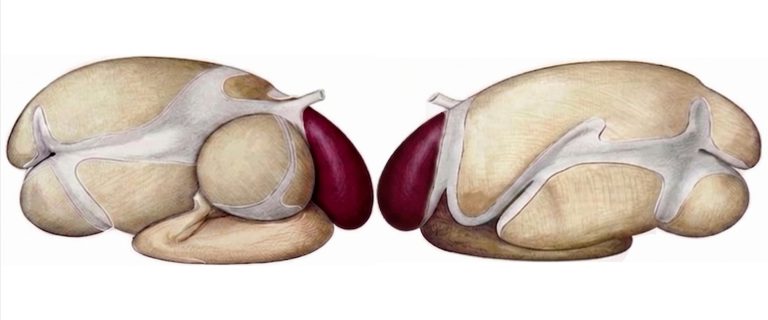
Figure 1. Lateral view of the forestomachs of a ruminant with emphasis on the reticulum (colored). Adapted from Popesko (1977).
2. Physiology of the Reticulum: Motility and Rumination
2.1. Biphasic Reticular contractions
The reticulum performs characteristic biphasic contractions: a first partial contraction (phase 1) followed by a more complete one (phase 2) that precede the primary rumen contraction, occurring approximately every minute. Through these contractions, the reticulum acts as a selective pump that discriminates particles according to their size and density: it retains large solid material and facilitates the passage of finely particulate material and liquid toward the omasum through the reticulo-omasal orifice (Mattioli, 2003; Pittroff & Kothmann, 2025).
2.2. Initiation of the Rumination Reflex
The reticulum plays a central role in triggering rumination. Stimulation of the mechanoreceptors in its wall by the presence of large solid material generates afferent impulses that reach the rumination center in the brainstem. The efferent response involves a sudden and forceful reticular contraction that, in synchrony with relaxation of the cardia and glottis, generates the negative pressure gradient necessary for bolus regurgitation. An adult bovine can devote 6–10 hours/day to rumination, with each cycle reducing the particle size of the forage and saturating it with bicarbonate saliva (Wainman, 1977; Mattioli, 2003).
2.3. The Esophageal Groove in Calves
In suckling calves, the esophageal groove, formed by two muscular lips on the dorsal wall of the reticulum, closes upon swallowing of milk or colostrum, redirecting the liquid directly to the abomasum and preventing its entry into the reticulo-rumen, whose microbiota is still developing. This reflex, mediated by the vagus nerve, gradually ceases with maturation (Porter, 1969; de Costa et al., 2007).
3. Impact of Mycotoxins on the Reticulum
3.1. Inflammation of the Reticular Mucosa
Chronic ingestion of contaminated feed causes local tissue inflammation in the reticular mucosa, with damage to the stratified squamous parakeratinized epithelium lining the characteristic hexagonal cells. Sustained irritation of the wall receptors predisposes to functional depression of the mechanoreceptors that regulate the characteristic biphasic contractions of the reticulum, slowing the transit of digesta toward the omasum (Fink-Gremmels, 2008; Gallo et al., 2015).
3.2. Neurotoxicity and Depression of Motility
Mycotoxins with neurotoxic activity, such as AFB1 and T-2 toxin, can accumulate in nerve tissue and compromise the vagal afferent pathways that control gastrointestinal motility. T-2 toxin exerts cytotoxic and immunotoxic effects that directly affect neurons of the reticular enteric plexus. As a consequence, coordinated contractions slow down, the rumination reflex is depressed, and digesta stagnates in the reticulo-rumen system, negatively impacting voluntary intake and motility of the entire anterior digestive tract (Mostrom & Jacobsen, 2020; Antonissen et al., 2014).
3.3. Suppression of Rumination and Cascade Consequences
The reduction in rumination time observed in cows exposed to diets with multiple mycotoxins has cascade consequences: it decreases saliva production and the supply of bicarbonate to the rumen, weakening the buffer system and predisposing the animal to subacute ruminal acidosis (SARA). Field studies in dairy cows fed diets naturally contaminated with DON, ZEN and fumonisins have recorded documented reductions in rumination time and energy-corrected milk production (ECM) compared with control cows (Gallo et al., 2023). This suppression of rumination, combined with stagnation of reticular transit, deteriorates the filtration exerted by the reticulo-omasal orifice and compromises the orderly flow of digesta toward the posterior compartments.
4. Clinical and Diagnostic Relevance
Mycotoxin-induced reticular dysfunction manifests as reduced rumination time (detectable by activity collars or ruminal sensors), reduced reticular contraction frequency (detectable by auscultation or ruminography), decreased voluntary intake without apparent infectious cause, and predisposition to frothy bloat due to unexpelled gas accumulation. In high-production herds, these signs are often misinterpreted as individual pathologies, when in reality they reflect a nutritional management problem with subclinical exposure to mycotoxins (Fink-Gremmels, 2008; Mostrom & Jacobsen, 2020).
Conclusion
The reticulum, although small in volume, is the engine of rumination and, therefore, of the salivary buffer system that sustains ruminal fermentation. Its alteration by mycotoxins, through inflammatory, neurotoxic, and motor depression mechanisms, triggers a chain of events that predisposes to SARA, dysbiosis and production loss.
To mitigate this impact, monitoring of rumination time must be complemented by the use of bioprotective mycotoxin mitigation strategies. These tools are essential to preserve the microbiota and epithelial integrity, representing the most effective line of action to protect the health and productivity of the ruminant.

