
Introducción
La demanda de productos pesqueros y acuícolas ha registrado un crecimiento sostenido desde 1961, con un incremento medio anual que se sitúa en el 3%. Este panorama global está liderado por el dinamismo del sector acuícola, cuyo volumen de producción ha superado de manera progresiva a los rendimientos de la pesca extractiva tradicional. Este desarrollo e impulso industrial responde directamente al incremento del consumo per cápita, el cual viene determinado por los procesos de urbanización, el aumento del poder adquisitivo y la evolución de las tendencias de consumo alimentario a escala mundial (FAO, 2022).
Dentro de este ámbito, la producción de crustáceos ocupa una fracción significativa de la actividad productiva total, con el camarón blanco (Litopenaeus vannamei) posicionándose como la cuarta especie acuícola más producida a nivel global y la segunda dentro del reino animal, precedida únicamente por la ostra del Pacífico (Crassostrea gigas) (FAO, 2022). El camarón blanco es un crustáceo decápodo marino originario de la costa oriental del Océano Pacífico, comúnmente asociado a aguas tropicales. Sin embargo, tolera un amplio rango de salinidades, así como variaciones moderadas de temperatura e incluso aguas continentales de baja salinidad. Dicha resiliencia, combinada con su rápido crecimiento y su excelente índice de conversión alimenticia, ha impulsado su posicionamiento como una especie clave para la producción acuática mundial.
A pesar de sus hábitos omnívoros, esta especie presenta requerimientos de proteína bruta moderadamente altos, los cuales suelen oscilar entre el 30% y el 40% de la composición total del alimento, dependiendo de la fase de desarrollo y la densidad de siembra del sistema productivo (Jobling et al., 2012). Tradicionalmente, el sector acuícola ha empleado la harina de pescado como la principal fuente proteica de alta calidad para cubrir dichas necesidades. No obstante, su volatilidad comercial y su limitado suministro global han impulsado una transición estratégica hacia fuentes de proteína vegetal más sostenibles y rentables, tales como los derivados de la soja y el maíz, complementados con aditivos funcionales que optimizan su digestibilidad.
Sin embargo, esta modificación en la formulación de los piensos acuícolas ha generado diversos desafíos entre los que destaca de manera prioritaria, un mayor riesgo de exposición de los peces a las micotoxinas.
Micotoxinas en acuicultura
Las micotoxinas son metabolitos secundarios tóxicos generados por diferentes especies de hongos, los cuales contaminan habitualmente tanto materias primas como alimentos terminados (Gruber-Dorninger et al., 2019). Su impacto se extiende a lo largo de toda la cadena alimentaria, y afecta al ser humano de manera directa, mediante la ingesta de productos vegetales contaminados, o indirecta, a través del consumo de alimentos de origen animal derivados de ganado expuesto a estas toxinas.
En el sector acuícola la exposición de las especies de cultivo no se restringe únicamente al consumo de alimento contaminado, ya que estos compuestos poseen la capacidad de persistir en el agua y en los sedimentos de los sistemas de producción. Esta diversidad en las vías de exposición no solo compromete severamente la salud y los índices productivos de los animales, sino que también pone en riesgo la inocuidad alimentaria del consumidor final debido a la potencial transferencia de residuos.
Desde la perspectiva del daño biológico, la toxicidad de estos compuestos desencadena un cuadro clínico complejo. Los principales efectos descritos incluyen el estrés oxidativo celular, lesiones histopatológicas severas en órganos diana como las branquias y el hepatopáncreas, alteraciones de conducta, caídas drásticas en la ganancia de peso y, en casos críticos, una elevada mortalidad. La gravedad de esta sintomatología es variable y depende estrechamente de la especie afectada, su fase de desarrollo, el tipo de micotoxina presente, la dosis ingerida y el periodo de exposición (Oliveira et al., 2020).
Por último, un factor crítico que agrava esta problemática es la alta termoestabilidad de las micotoxinas. Esta resistencia estructural les permite soportar de forma eficaz los tratamientos térmicos y los procesos de extrusión habituales en las plantas de fabricación de piensos acuícolas, permaneciendo activas en el alimento final (Gbashi et al., 2019; Sueck et al., 2019).
Micotoxinas en camarón blanco
A continuación, se detallan los efectos fisiopatológicos de las micotoxinas más estudiadas en el camarón blanco. No obstante, cabe destacar que, más allá de la acción individual de estos compuestos, el escenario más frecuente en la naturaleza es la co-contaminación por múltiples micotoxinas en lugar de su detección aislada. La relevancia de este fenómeno radica en que estas toxinas suelen presentar sinergismo, de manera que la presencia simultánea de varias en un mismo alimento potencia su toxicidad.
Aflatoxina B1
Las aflatoxinas son un conjunto de micotoxinas producidas por hongos del género Aspergillus, principalmente A. flavus y A. parasiticus, e incluyen variantes B1, B2, G1 y G2. De todas ellas, la aflatoxina B1 (AFB1) destaca por su elevado poder carcinógeno y su tendencia a bioacumularse en órganos diana, tales como el hepatopáncreas y en el tejido muscular.
En el cultivo de camarón blanco, la exposición a AFB1 desencadena una cascada de efectos fisiopatológicos que comprometen severamente los parámetros productivos. Investigaciones como las de Yu et al. (2018) demostraron que inclusiones desde 500 ppb en la dieta inducen un descenso en el crecimiento de hasta un 39,44% y una caída del 11,25% de la supervivencia en comparación con animales alimentados con dietas libres de micotoxinas; estos efectos se agudizan con dosis más elevadas. Este impacto negativo en el rendimiento se asocia a disfunciones digestivas, alteraciones de la microbiota, estrés oxidativo, daño histológico en el hepatopáncreas e inmunosupresión.
Respecto a la disfunción gastrointestinal, Su et al. (2021) reportaron una disminución drástica en la actividad de enzimas digestivas clave (proteasa, lipasa y amilasa) a partir de dosis de 2500 ppb. De forma similar, bajo condiciones de estrés por concentraciones de 5000 ppb, Wang et al. (2018) evidenciaron un colapso de la microbiota intestinal. Este desequilibrio se caracterizó por el descenso de grupos bacterianos benéficos como Flavobacterium y Bacteroidetes, y el incremento colateral de patógenos potenciales como Vibrio y Photobacterium.
En lo referente al estrés oxidativo, el mismo estudio de Wang et al. (2018) constató un incremento inicial en la actividad de las enzimas de defensa antioxidante, tales como la superóxido dismutasa (SOD), la catalasa (CAT) y la glutatión peroxidasa (GPX), a partir de 5000 ppb, seguido de un colapso total del sistema defensivo tras dos semanas de exposición. Esta caída se vio acompañada por un aumento del malondialdehído (MDA), un biomarcador clave de la peroxidación lipídica y el daño celular.
Este escenario patológico se vincula estrechamente con alteraciones estructurales en el hepatopáncreas. Su et al. (2025) reportaron la ruptura de células epiteliales y un incremento del lumen a partir de contaminaciones de 2500 ppb en el alimento. Así mismo, este daño hepático se refleja en el plasma mediante el incremento de biomarcadores de ruptura celular como la alanina aminotransferasa (ALT) y la aspartato aminotransferasa (AST), de acuerdo con lo observado por Yu et al. (2018).
Por último, la inmunosupresión provocada por esta micotoxina incrementa la susceptibilidad a agentes patógenos. Este compromiso inmunológico se manifiesta a través de una respuesta inflamatoria exacerbada, reflejada en el incremento plasmático de la óxido nítrico sintasa inducible (iNOS), y una notable reducción en la expresión de genes esenciales para la inmunomodulación, tales como los pertenecientes a las vías Toll y Dorsal.
Fumonisinas
Las fumonisinas son micotoxinas originadas por hongos del género Fusarium, principalmente por las especies F. verticillioides y F. proliferatum, las cuales son responsables de la síntesis de las variantes B1, B2 y B3 (Cáceres et al., 2020).
En el cultivo de camarón blanco, de forma similar a lo observado con la AFB1, la exposición a la fumonisina B1 (FB1) compromete severamente los parámetros zootécnicos. Investigaciones como las de Kracizy et al. (2021) demostraron que inclusiones desde 1715 ppb en la dieta inducen una reducción del 19,91% en la ganancia de peso, en comparación con animales alimentados con dietas libres de contaminantes.
Además de afectar el crecimiento, esta toxina altera la calidad final del producto mediante la desestabilización de las proteínas musculares; García-Morales et al. (2015) reportaron reducciones en la proteína soluble del músculo miofibrilar y una menor capacidad de retención de agua a partir de dosis de 200 ppb.
Este impacto negativo se agudiza al alcanzar concentraciones de 1000 ppb, presentándose alteraciones severas como la hidrólisis de las cadenas de miosina (Mexía-Salazar et al., 2008), lo que reduce significativamente la vida útil del producto durante el almacenamiento.
Respecto al daño tisular, se han registrado diversos deterioros histopatológicos en órganos diana como el hepatopáncreas y las branquias. Mexía-Salazar et al. (2008) evidenciaron que contaminaciones de 500 ppb desencadenan la deformación y vacuolización de los túbulos hepatopancreáticos, así como signos iniciales de melanización. Al incrementarse la exposición a dosis de 1000 ppb, el daño progresa hacia una marcada infiltración de hemocitos y necrosis tisular. En este mismo nivel de inclusión (1000 ppb), los autores constataron que el impacto patológico se extiende a las branquias, provocando atrofia y graves daños anatómicos.
Por último, el compromiso del sistema inmune se manifiesta de forma drástica a partir de inclusiones de 500 ppb. De acuerdo con Mexía-Salazar et al. (2008), esta concentración induce una reducción severa en el conteo total de hemocitos circulantes y disminuye la tasa de producción del anión superóxido. Esto deprime la actividad fagocítica, un mecanismo clave en la defensa frente a patógenos. Asimismo, se observa una supresión de la inmunidad humoral en este mismo umbral de dosis, reflejada en la caída de la actividad de enzimas inmunológicas clave como la fenoloxidasa (PO).
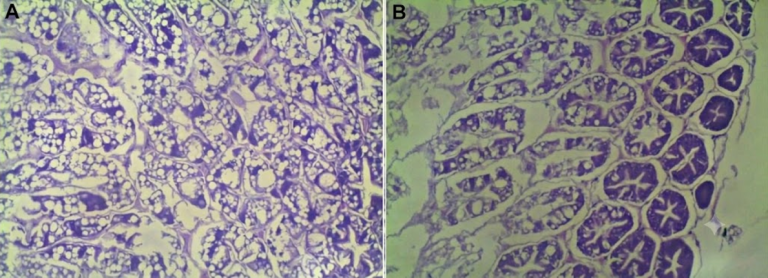
Figura 1. Cortes histológicos de hepatopáncreas de camarón blanco. (A) Tejido íntegro; (B) Atrofia celular y vacuolización por acción de FB1 (Kracizy et al., 2021).
Ocratoxina A
Las ocratoxinas constituyen un grupo de metabolitos fúngicos generados principalmente por especies de los géneros Aspergillus y Penicillium. Dentro de este grupo, la ocratoxina A (OTA) destaca como la variante más prevalente y sobre la cual se han registrado más efectos en especies de producción acuícola, incluyendo el camarón.
De acuerdo con las investigaciones de Albuquerque et al. (2016), la exposición a dosis a partir de 500 ppb de OTA es suficiente para inducir caídas significativas en el rendimiento zootécnico. Este impacto negativo muestra una correlación directa con el aumento de la concentración de la micotoxina en el alimento: A dosis de 500 ppb, se observa una reducción del 6 % en la ganancia media de peso en comparación con los animales sanos, así como un deterioro en el índice de conversión alimenticia, el cual se incrementa de 1,01 a 1,09. A dosis a 1000 ppb, el índice de conversión empeora drásticamente hasta alcanzar un valor de 1,23, mientras que la tasa de crecimiento experimenta una contracción del 17,82 % respecto al lote de control.
Paralelamente, dicho estudio constató un preocupante fenómeno de transferencia (carry-over) de la toxina hacia los tejidos destinados al consumo humano. Específicamente, la inclusión de apenas 100 ppb de OTA en el alimento dio lugar a la retención de residuos de 0,20 ppb en la musculatura abdominal de los animales. Esta acumulación tisular no solo compromete seriamente la viabilidad económica y productiva del lote, sino que también introduce un riesgo directo para la seguridad alimentaria y la salud del consumidor final.
Toxina T2
La toxina T2 es un tricoteceno producido por hongos del género Fusarium. Estos patógenos colonizan cultivos de maíz, trigo y soja, lo que resulta en la contaminación de los granos que posteriormente se emplean en la elaboración de piensos acuícolas (Schatzmayr y Streit, 2013).
De acuerdo con los hallazgos de Qiu et al. (2016), la exposición a una dosis baja de apenas 500 ppb es suficiente para comprometer de forma drástica la productividad en las granjas. Al evaluar dietas contaminadas con 500 ppb de toxina T2 frente a controles libres de micotoxinas, se observaron los siguientes efectos: Disminución de la tasa de supervivencia desde un 92,22% en hasta un 78,88%; la ganancia de peso se ralentizó significativamente, descendiendo del 22% al 18,30%.
Este declive en el rendimiento productivo está estrechamente vinculado a un efecto inmunosupresor severo, caracterizado por una caída en la actividad de la fenoloxidasa (PO), cuya absorbancia disminuye desde 17,93 min-1×mg de proteína-1 en animales sanos, hasta 12,09 y 9,61 con dosis de 500 y 1200 ppb del compuesto, respectivamente. Además, se genera una reducción drástica en el recuento de hemocitos totales, el cual cae de 8,00×106 a 5,00×106 al alcanzar las 2400 ppb, dejando al organismo desprotegido frente a patógenos secundarios.
A nivel fisiológico, el hepatopáncreas actúa como el principal órgano diana de la toxicidad, manifestando alteraciones que evidencian el colapso metabólico del sistema digestivo.
La evaluación enzimática realizada por Qiu et al. (2016) demuestra un desplome en la actividad de la transaminasa glutámico-pirúvica (GPT), la cual se reduce de 62,73 U/L en dieta control a 25,03 U/L con una dosis de 1200 ppb de toxina T2. En consonancia con este biomarcador de daño tisular, los análisis histopatológicos integrados de Qiu et al. (2016) y Bi et al. (2019) confirman una progresión de lesiones dosis-dependiente en la microestructura hepatopancreática: A 500 ppb cursan con procesos inflamatorios, atrofia celular y una coloración rojiza macroscópica. A partir de 1200 ppb y 1500 ppb el cuadro evoluciona hacia una marcada vacuolización celular; y en concentraciones más elevadas culmina en lisis celular generalizada y necrosis severa del epitelio tubular.
De forma simultánea, la integridad cualitativa del tejido muscular y el perfil lipídico del camarón sufren un deterioro sistémico que deprecia el valor del producto final. En un diseño experimental de 20 días con ejemplares de 4 g, Bi et al. (2019) determinaron que la ingesta de toxina T2 altera el metabolismo de los lípidos corporales, provocando una reducción inicial en el contenido total de ácidos grasos saturados (SFA), monoinsaturados (MUFA) y poliinsaturados (PUFA) en el músculo bajo condiciones de dosificación baja.
Esta desnutrición química se correlaciona directamente con el daño en la carne del crustáceo, en donde la microscopía revela una mayor separación miofibrilar en dosis de hasta 1500 ppb; un cuadro patológico que se agrava exponencialmente a partir de las 4500 ppb y hasta las 13500 ppb mediante la pérdida de continuidad del tejido, la separación de los sarcómeros y la pérdida de la estructura compacta del músculo, afectando negativamente la consistencia y la calidad de la biomasa cosechada.
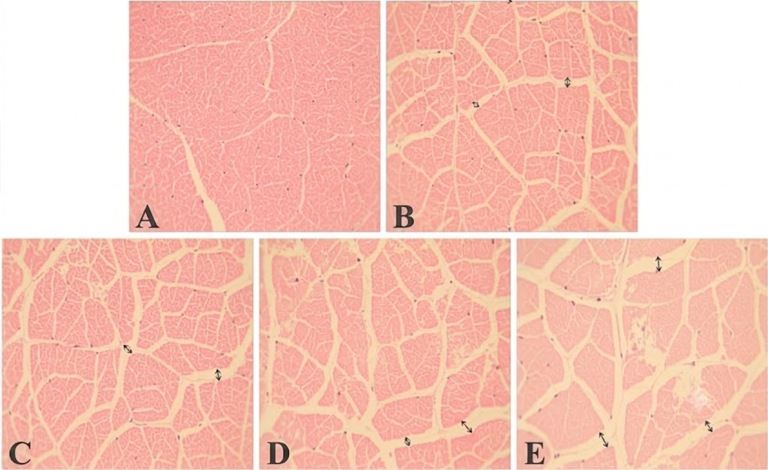
Figura 2. Cortes histológicos de músculo de camarón con dosis crecientes de toxina T2: (A) 0 ppb; (B) 500 ppb; (C) 1500 ppb; (D) 4500 ppb; (E) 13500 ppb. Separación miofibrilar y de sarcómeros creciente (Bi et al., 2019).
Deoxinivalenol
El deoxivalenol (DON) es otro tricoteceno de gran relevancia generado también por hongos del género Fusarium. Al colonizar cultivos agrícolas como el maíz y el trigo, contamina las materias primas presentes en los piensos empleados en la cría de camarón.
Según la investigación desarrollada por Xie et al. (2018), la ingesta de deoxinivalenol induce una penalización progresiva en los índices zootécnicos del crustáceo. Los resultados demuestran una reducción ligera en la ganancia de peso a partir de una inclusión baja de 250 ppb, un efecto que se agrava significativamente al alcanzar concentraciones más elevadas. Esta merma en el rendimiento productivo es el reflejo de una severa disfunción metabólica y del coste energético que asume el animal para intentar contrarrestar la toxicidad en su organismo.
A nivel tisular, el deoxinivalenol ejerce una acción dependiente de la dosis que desestabiliza las barreras digestivas y funcionales del camarón, localizándose el daño principalmente en el hepatopáncreas y el tracto intestinal. Las evaluaciones histológicas de Xie et al. (2018) revelan que dosis iniciales de 250 ppb provocan hipertrofia en las células B del hepatopáncreas, una respuesta celular defensiva que evoluciona negativamente al incrementarse la toxina a 500 ppb, donde se evidencia la fusión de los pliegues de la mucosa intestinal, un aumento en el diámetro de los túbulos hepatopancreáticos y la aparición de apoptosis celular. Al alcanzar la concentración crítica de 1000 ppb, la patología microscópica revela un escenario de apoptosis avanzada, caracterizado por la presencia masiva de cuerpos apoptóticos y una notable hinchazón del retículo endoplasmático, lo que confirma el colapso de la maquinaria celular encargada de la síntesis de proteínas.
De acuerdo con lo expuesto por Xi et al. (2008), los daños en los tejidos están directamente vinculados al estrés oxidativo y a la inmunosupresión. La respuesta fisiológica ante la toxina depende estrictamente del nivel de exposición. A partir de las 500 ppb se activan los mecanismos de desintoxicación celular mediante el incremento de la enzima glutatión S-transferasa (GST) y una mayor expresión de la superóxido dismutasa (SOD) y la glutatión peroxidasa (GPX).
Sin embargo, el sistema inmune muestra un comportamiento diferenciado según la dosis. Mientras que una exposición baja, de 250 ppb, actúa como un estresor agudo que sobreestimula la vía NF-kB y la proteína HSP70 para reparar el daño celular, las dosis elevadas de 1.000 ppb provocan el efecto contrario. Ante un desafío sostenido, este mecanismo de defensa colapsa, derivando en una inmunosupresión sistémica y la sobre activación del sistema profenoloxidasa (proPO).
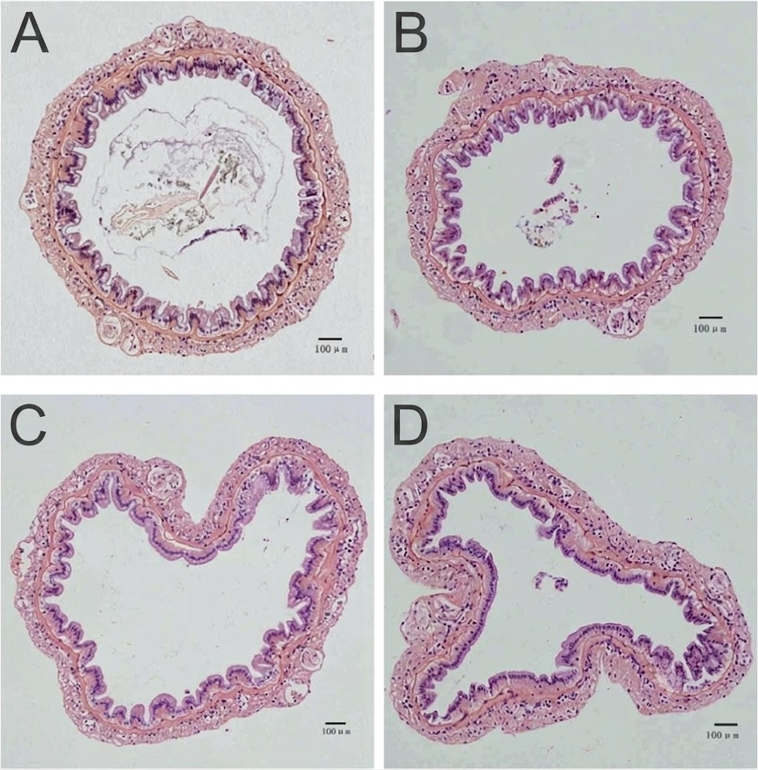
Figura 3. Cortes histológicos de intestino: (A) Tejido íntegro; (B, C, D) Alteraciones estructurales y de la mucosa ocasionadas por dosis crecientes de DON (Xie et al., 2018).
Micotoxinas en otros crustáceos
Si bien el camarón blanco suele concentrar la mayor parte de la atención de la investigación acuícola debido a su volumen de producción, la sensibilidad a las micotoxinas no es exclusiva de esta especie. El impacto de estos metabolitos fúngicos se extiende a otros crustáceos de alto valor comercial, tanto marinos como de agua dulce, manifestándose a través de alteraciones metabólicas, inmunosupresión y estrés oxidativo.
Camarón tigre
El camarón tigre (Penaeus monodon) es un pilar acuícola global gracias a su gran tamaño, rápido crecimiento y alto valor. Aunque su producción sufrió un fuerte declive por crisis sanitarias en comparación con el camarón blanco, hoy experimenta un resurgimiento gracias al desarrollo de líneas genéticas libres de patógenos. De este modo, se mantiene como un motor económico clave y una fuente masiva de divisas en regiones tropicales.
En el contexto de las micotoxinas, Supamattaya et al. (2005) reportaron que la exposición prolongada a la OTA y el DON durante un periodo de ocho semanas desencadena alteraciones fisiológicas específicas en esta especie, afectando de manera diferenciada su rendimiento defensivo y metabólico según la dosis administrada.
Al evaluar los efectos de la OTA en concentraciones de 100, 200 y 1000 ppb, los impactos más críticos se manifestaron con la dosis más elevada (1000 ppb), provocando: Una disminución significativa en la actividad de la enzima fenoloxidasa (PO), lo que compromete el sistema inmune del crustáceo; y una ligera reducción en la ganancia de peso corporal de los animales.
Por el contrario, el DON mostró un comportamiento marcadamente distinto al ser probado en dosis de 500, 1000 y 2000 ppb, ya que no alteró la actividad de la fenoloxidasa ni el conteo total de hemocitos.
A pesar de estas diferencias y de la ausencia de lesiones histopatológicas en tejidos clave como las branquias, el tejido nervioso o el hepatopáncreas, ambas micotoxinas compartieron un impacto sistémico negativo. Supamattaya et al. (2005) identificaron una reducción generalizada en los niveles plasmáticos de fosfatasa alcalina (ALP), transaminasa glutámico-oxaloacética (GOT) y transaminasa glutámico-pirúvica (GPT). La caída de estas enzimas hepáticas e inmunofisiológicas constituye un indicador indirecto crucial de que tanto la OTA como el DON son capaces de alterar y deteriorar el correcto funcionamiento celular del hepatopáncreas.
Cangrejo chino
El cangrejo chino (Eriocheir sinensis), es una de las especies de agua dulce más valiosas de Asia Oriental, caracterizado por una fuerte demanda tradicional y de lujo gastronómico. Su cultivo se realiza de forma masiva en lagos y estanques de China mediante sistemas seminaturales de propagación artificial.
En el ámbito de su crianza, el análisis de la contaminación de los piensos por micotoxinas es un factor crítico no solo para la productividad, sino para la seguridad alimentaria, dado el riesgo potencial que representa la acumulación de toxinas en su carne comestible para los consumidores humanos. Investigaciones recientes lideradas por Yang et al. (2023) y Qiu et al. (2016) han arrojado luz sobre los severos efectos fisiológicos que provocan tanto la toxina T2 como la AFB1 en esta especie, evidenciando dinámicas de toxicidad crónicas y agudas altamente perjudiciales.
Al evaluar la toxina T2 administrada de forma crónica durante 56 días en cangrejos juveniles (en dosis de 600, 2500 y 5000 ppb de alimento), se demostró un impacto directo en los índices productivos, reduciendo significativamente el crecimiento y la ganancia de peso en todas las concentraciones evaluadas, además de incrementar la mortalidad en la dosis más alta (5000 ppb). A nivel fisiológico, esta sustancia desencadenó un cuadro severo de estrés oxidativo, caracterizado por la elevación del malondialdehído (MDA) y el desplome de las defensas antioxidantes superóxido dismutasa (SOD) y la glutatión peroxidasa (GPX), combinado con una profunda inmunosupresión manifestada en la caída del recuento de hemocitos y del estallido respiratorio. Asimismo, se constató la inducción de procesos inflamatorios y de apoptosis desde la dosis más baja, acompañados de daños morfológicos severos en el hepatopáncreas que destruyeron la membrana basal de sus túbulos y alteraron el sistema de desintoxicación celular.
Por otro lado, la evaluación del impacto agudo de la AFB1 mediante una inyección única de 400 µL (60 mg/L) reveló una toxicidad inmediata e intensa concentrada en el hepatopáncreas a los 30 y 60 minutos postexposición. Este daño activó la expresión del gen de la sorbitol deshidrogenasa, un marcador directo de lesión celular en este tejido, provocando una fuerte alteración inmunitaria y de estrés oxidativo.
Cangrejo rojo americano
El cangrejo rojo americano (Procambarus clarkii), originario de Norteamérica, es la especie de cangrejo de río más cultivada del mundo debido a su notable resistencia, rápido crecimiento y alta fertilidad. Su producción lidera la acuicultura de agua dulce en países como China y Estados Unidos, integrándose con éxito en sistemas de rotación de cultivos como el arroz.
En el marco de la seguridad e inmunidad de esta especie, un estudio desarrollado por Wen et al. (2023) evaluó los efectos de la exposición al DON mediante la administración de una dosis de 3000 ppb. El tratamiento generó una respuesta fisiológica notable a nivel defensivo, registrándose un incremento metabólico en el suero de las enzimas antioxidantes superóxido dismutasa (SOD), catalasa (CAT) y glutatión S-transferasa (GST), así como un aumento en la actividad de la enzima inmunitaria fosfatasa alcalina (AKP).
Conclusión
El cambio estratégico hacia proteínas vegetales en los piensos acuícolas ha incrementado la exposición del camarón blanco a las micotoxinas. Como se ha detallado, la presencia de toxinas como AFB1, FB1, OTA, T2 y DON desencadena un colapso multiorgánico caracterizado por estrés oxidativo, inmunosupresión, graves daños histológicos en el hepatopáncreas y pérdida de la calidad muscular, lo que merma directamente el rendimiento productivo de las explotaciones.
Por ello, la gestión rigurosa de las micotoxinas mediante el uso de soluciones biotecnológicas eficientes es un pilar imprescindible para proteger la salud de los animales, optimizar los parámetros zootécnicos y garantizar los más altos estándares de inocuidad exigidos para el consumo humano.


