
Impacto de las micotoxinas en los compartimentos gástricos del rumiante
Introducción
El rumen es mucho más que un simple depósito de fermentación: es un ecosistema microbiano de extraordinaria complejidad funcional, comparable en diversidad a los microbiomas más estudiados de la biología moderna. En un bovino adulto, cada mililitro de contenido ruminal aloja entre 10¹⁰ y 10¹¹ bacterias, 10⁵ a 10⁶ protozoos ciliados, 10³ a 10⁵ hongos anaeróbicos y una comunidad de arqueas metanogénicas que producen metano como subproducto de la fermentación. Esta comunidad, organizada en una red trófica de altísima precisión, convierte la biomasa vegetal lignocelulósica indigerible para los mamíferos monogástricos en ácidos grasos volátiles (AGV) y proteína microbiana, las dos fuentes energéticas y proteicas dominantes del rumiante (Annison & Bryden, 1998; McAllister et al., 2025).
Esta complejidad, sin embargo, es también una vulnerabilidad. El ecosistema ruminal requiere condiciones físico-químicas estrictamente reguladas: temperatura de 38-42 °C, potencial redox marcadamente negativo (-250 a -450 mV), anaerobiosis estricta y pH entre 5,5 y 7,0. Cualquier perturbación de estas condiciones puede desencadenar una disbiosis, un desequilibrio cualitativo y cuantitativo de la microbiota con consecuencias en cascada sobre la eficiencia digestiva, la salud del epitelio y el estado metabólico general del animal (Vivares et al., 2025; Motyka et al., 2025).
Las micotoxinas, metabolitos secundarios producidos por hongos filamentosos de los géneros Aspergillus, Fusarium, Penicillium y Alternaria, representan uno de los agentes perturbadores del ecosistema ruminal más frecuentes y subestimados en la práctica clínica y nutricional. A diferencia de otras noxas dietéticas, las micotoxinas no actúan únicamente como tóxicos sistémicos: su primera y a menudo más devastadora diana es el propio microbioma ruminal. Este artículo analiza los mecanismos específicos por los que las principales micotoxinas, aflatoxinas (AFs), deoxinivalenol (DON), zearalenona (ZEA), fumonisinas (FBs) y toxina T-2/HT-2 alteran la fisiología del rumen, con especial énfasis en la disbiosis, la fermentación y la permeabilidad epitelial.
1. Anatomía e histología del rumen: bases para entender el daño por micotoxinas
El rumen ocupa la mayor parte del hemicuerpo izquierdo de la cavidad abdominal, con una capacidad de 100-250 litros en bovinos adultos. Sus cinco sacos (dorsal, ventral, craneal y los ciegos caudodorsal y caudoventral), delimitados por robustos pilares musculares como se observa en la Figura 1, no son meras divisiones anatómicas: cada uno cumple un rol específico en la estratificación y el movimiento del contenido. La cúpula de gas, el estrato de fibra flotante, el estrato fangoso en fermentación activa y el estrato líquido ventral se organizan por diferencias de densidad y son redistribuidos por las contracciones primarias y secundarias del ciclo retículo-ruminal (Mattioli, 2003; Yu et al., 2026).
Histológicamente, la mucosa ruminal está revestida por un epitelio plano estratificado escamoso paraqueratinizado, sin glándulas secretoras. Su función es exclusivamente absortiva. Las papilas digitiformes y linguiformes que recubren toda la superficie interna pueden incrementar el área de absorción total hasta 1,0-1,5 m² en bovinos adultos con dieta mixta, y hasta un 400% más en animales con dietas altamente fermentables. El butirato, uno de los tres AGV principales actúa como el principal agente trófico sobre este epitelio: estimula la proliferación de las células del estrato basal y mantiene la arquitectura papilar. La exposición a micotoxinas que deprimen la producción de butirato tiene, por tanto, consecuencias directas sobre la integridad morfológica de la mucosa absortiva (de Costa et al., 2007; Vivares et al., 2025).
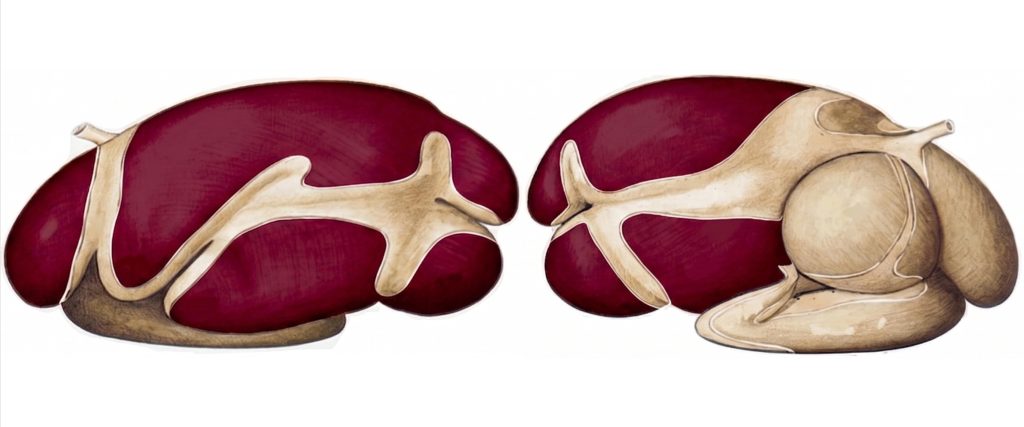
Figura 1. Vista lateral de los preestómagos de un rumiante con énfasis en el rumen (coloreado). Adaptado de Popesko (1977).
2. Fisiología de la fermentación ruminal
2.1. El ecosistema microbiano
Las bacterias celulolíticas (Fibrobacter succinogenes, Ruminococcus albus, Ruminococcus flavefaciens) atacan físicamente la fibra mediante celulasomas, produciendo endoglucanasas y exoglucanasas que hidrolizan los enlaces β-1,4-glucosídicos de la celulosa. Los hongos anaeróbicos (Neocallimastix, Piromyces, Orpinomyces) actúan como pioneros en la colonización de la fibra: sus rizoides penetran físicamente la pared vegetal, aumentando la superficie de ataque bacteriano. Los protozoarios ciliados (Entodinium, Diplodinium) modulan la dinámica microbiana y actúan como reservorio de AGV durante las fluctuaciones de pH.
Los productos finales de la fermentación son los AGV (acetato, propionato y butirato), CO₂ y CH₄. Las proporciones varían con la dieta: forrajes generan 65:20:15 (acetato:propionato:butirato), mientras que los concentrados ricos en almidón desplazan la fermentación hacia el propionato (55:30:15). El acetato es el substrato primario para la síntesis de grasa láctea; el propionato, la principal fuente de carbono para la gluconeogénesis hepática; y el butirato, el sustrato energético del propio epitelio ruminal (Annison & Bryden, 1999).
2.2. Absorción de AGV y tamponamiento
La absorción de AGV ocurre principalmente por difusión pasiva de las formas no disociadas a través de la membrana apical del epitelio. La fracción disociada se absorbe mediante intercambiadores SCFA⁻/HCO₃⁻, generando un flujo neto de bicarbonato hacia la luz ruminal que tamponea el pH. El butirato es metabolizado en >90% por los propios colonocitos, convirtiéndose en β-hidroxibutirato y acetoacetato antes de ser liberado a la circulación portal. El acetato y el propionato fluyen directamente al hígado (Annison & Bryden, 1998).
El sistema tampón del rumen depende críticamente de la saliva: un bovino adulto secreta 100-200 litros/día con 120-130 mEq/L de bicarbonato. La rumia, que puede ocupar 6-10 horas/día, estimula la salivación. Cualquier agente que deprima el tiempo de rumia reduce directamente el aporte de tampón al rumen, predisponiendo a acidosis ruminal sub-aguda (SARA).
3. Impacto de las micotoxinas sobre el rumen
3.1. Límites de la detoxificación ruminal
El rumen constituye la primera línea de defensa del rumiante frente a las micotoxinas ingeridas, pero esta capacidad de detoxificación ruminal tiene límites biológicos bien definidos. La microbiota ruminal puede degradar ciertos metabolitos fúngicos: la ocratoxina A (OTA) es convertida en ocratoxina alfa (OTα), de menor toxicidad; algunos tricotecenos como el diacetoxiscirpenol (DAS) son transformados moderadamente por los protozoos. Sin embargo, la aflatoxina B1 (AFB1) y el deoxinivalenol (DON) resisten prácticamente sin modificación al ambiente ruminal, manteniendo su potencial tóxico intacto (Debevere et al., 2020; Gallo et al., 2024).
En esta misma línea, las fumonisinas (FBs) muestran una degradación ruminal mínima (inferior al 10%), lo que permite que la totalidad de la toxina alcance el intestino delgado sin perder su actividad biológica (Krüger et al, 2022; Gallo et al., 2024). Por otro lado, la toxina T-2 presenta un escenario mixto: aunque los protozoos ciliados pueden realizar una metabolización moderada hacia metabolitos menos tóxicos, esta capacidad se satura rápidamente ante niveles de contaminación elevados, permitiendo el paso de la toxina intacta a los compartimentos posteriores (Debevere et al., 2020).
Un caso especialmente preocupante es el de la zearalenona (ZEA). La microbiota ruminal la convierte principalmente en α-zearalenol (α-ZEL), un metabolito con una potencia estrogénica notablemente superior a la molécula original. En lugar de detoxificar, el rumen metaboliza el compuesto, exacerbando los trastornos reproductivos. Estudios en vacas Holstein con dietas ricas en forraje demostraron que la ZEA y sus metabolitos alteran rápidamente el microbioma ruminal desde las primeras 48 horas de exposición (Hartinger et al., 2023).
3.2. Disbiosis inducida: impacto sobre el microbioma
Las micotoxinas que superan o eluden la barrera ruminal ejercen efectos antimicrobianos directos sobre la comunidad residente. La exposición a FBs y la ZEA reduce significativamente las poblaciones de Lachnospiraceae y Prevotellaceae, dos familias bacterianas clave en la fermentación de fibra y en la producción de propionato, respectivamente. La ZEA reduce el pH ruminal y la concentración total de ácidos grasos de cadena corta (AGCC), comprometiendo la disponibilidad energética del animal. Las FBs alteran la diversidad β del microbioma y las funciones metagenómicas predichas del consorcio microbiano, documentadas con solo 2 días de exposición (Krüger et al., 2022).
La depresión de las bacterias celulolíticas por las micotoxinas deteriora la degradación de la fibra y reduce drásticamente la síntesis de proteína microbiana. La producción de AGV cae, generando un déficit energético que el animal no puede compensar por incremento del consumo voluntario, el cual también se ve deprimido. La AFB1 reduce la producción de gas, la digestibilidad de la materia seca y las concentraciones de nitrógeno-amoniacal en el rumen, evidenciando un deterioro global de la fermentación (Kemboi et al., 2020). En sistemas de alta producción, la cinética de paso acelerada reduce el tiempo de retención ruminal, limitando aún más la capacidad de detoxificación y permitiendo que mayores proporciones de toxinas pasen intactas a los demás compartimentos (Gallo et al., 2024).
Este colapso microbiológico y la alteración del pH resultante derivan en lesiones inflamatorias visibles en la mucosa, como se observa en la Imagen 1.
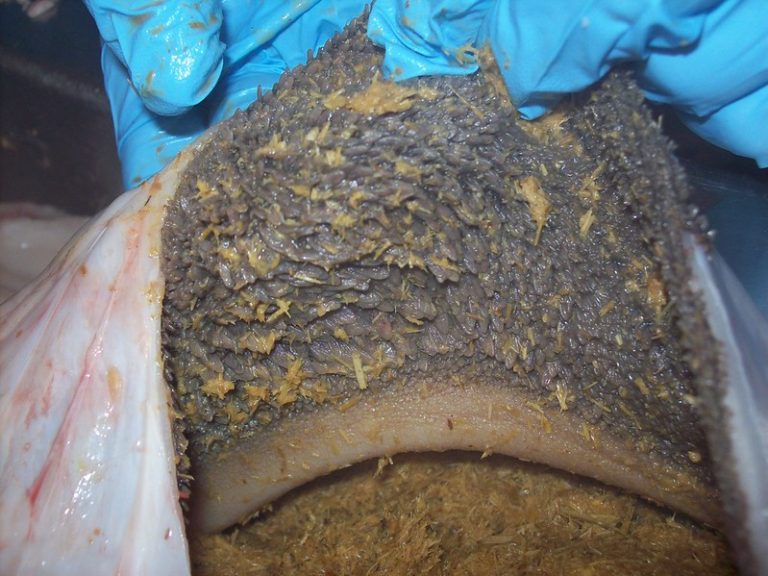
Imagen 1. Mucosa ruminal con signos evidentes de ruminitis. Se observa una marcada inflamación del epitelio y alteración de las papilas ruminales, lo que compromete la capacidad absortiva de AGV.
3.3. Permeabilidad epitelial y síndrome de intestino permeable
La disbiosis inducida por micotoxinas también incrementa la permeabilidad del epitelio ruminal. El aumento de la permeabilidad de la mucosa del rumen (Imagen 1), junto con el daño directo al epitelio gastrointestinal, son marcadores frecuentes de la micotoxicosis (Antonissen et al., 2014; Gao et al., 2020). La destrucción de las uniones estrechas (tight junctions) facilita la translocación de lipopolisacáridos (LPS) bacterianos y de las propias toxinas hacia la circulación portal, desencadenando inflamación sistémica y sobrecargando la capacidad detoxificadora hepática. Como se detalla en la Imagen 2, este proceso rompe la barrera defensiva natural, permitiendo el paso de agentes nocivos por vía paracelular.
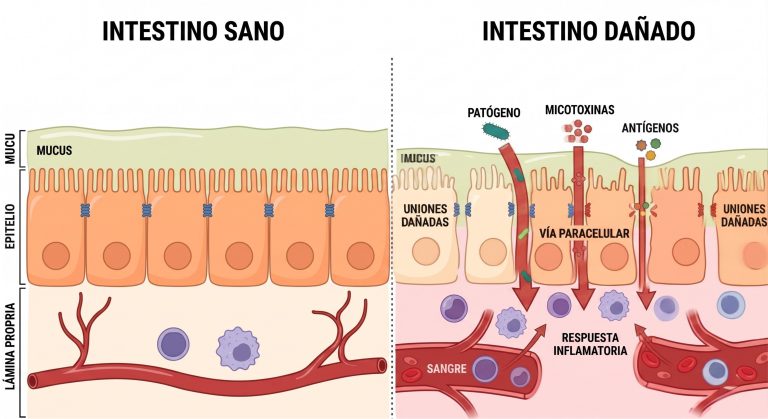
Imagen 2. Representación esquemática de la pérdida de integridad en las uniones estrechas. A la izquierda, un epitelio sano con barrera funcional; a la derecha, un epitelio dañado donde la vía paracelular permite la traslocación de lipopolisacáridos (LPS) y toxinas hacia la circulación sanguínea.
4. Consecuencias productivas y clínicas
La integración de los mecanismos anteriores produce un síndrome metabólico-digestivo que se manifiesta clínicamente como: reducción del consumo voluntario, descenso de la producción de leche y de la ganancia diaria de peso, peor eficiencia de conversión del alimento, mayor incidencia de SARA y sus complicaciones (laminitis, hepatitis, abscesos hepáticos) y predisposición a enfermedades infecciosas por inmunosupresión. En vacas en transición, la concurrencia de balance energético negativo y disbiosis micotoxicógena eleva exponencialmente el riesgo de cetosis, retención de placenta y metritis (Donkor et al., 2016; Motyka et al., 2025).
Conclusión
El rumen es la primera y más importante diana de las micotoxinas en el rumiante. Su microbioma, extraordinariamente sensible a estas toxinas, puede ser perturbada incluso con exposiciones cortas y a concentraciones subcrónicas. La detoxificación ruminal es real pero limitada, y en el caso de la ZEA puede incluso generar metabolitos de mayor potencia biológica. La gestión nutricional preventiva, monitoreo de micotoxinas en materias primas, uso de estrategias de bioprotección avanzadas para la mitigación de micotoxinas y preservación de la diversidad del microbioma es la estrategia más coherente con el conocimiento científico actual para proteger la salud y la productividad del rumiante de alta producción.


