
Impact of mycotoxins on the gastric compartments of the ruminant
Introduction
The rumen is more than a simple fermentation vessel: it is a microbial ecosystem of extraordinary functional complexity, comparable in diversity to the most studied microbiomes in modern biology. In an adult bovine, each milliliter of ruminal content harbors between 10¹⁰ and 10¹¹ bacteria, 10⁵ to 10⁶ ciliated protozoa, 10³ to 10⁵ anaerobic fungi and a community of methanogenic archaea that produce methane as a byproduct of fermentation. This community, organized in a highly precise trophic network, converts the lignocellulosic plant biomass, indigestible for monogastric mammals into volatile fatty acids (VFA) and microbial protein, the two dominant energy and protein sources of the ruminant (Annison & Bryden, 1998; McAllister et al., 2025).
This complexity, however, is also a vulnerability. The ruminal ecosystem requires strictly regulated physicochemical conditions: temperature of 38–42 °C, markedly negative redox potential (−250 to −450 mV), strict anaerobiosis and pH between 5.5 and 7.0. Any disruption of these conditions can trigger dysbiosis, a qualitative and quantitative imbalance of the microbiota with cascade consequences for digestive efficiency, epithelial health and the general metabolic status of the animal (Vivares et al., 2025; Motyka et al., 2025).
Mycotoxins, secondary metabolites produced by filamentous fungi of the genera Aspergillus, Fusarium, Penicillium and Alternaria, represent one of the most frequent and underestimated disruptive agents of the ruminal ecosystem in clinical and nutritional practice. Unlike other dietary noxa, mycotoxins do not act solely as systemic toxins: their first and often most devastating target is the ruminal microbiome itself. This article analyzes the specific mechanisms by which the main mycotoxins, aflatoxins (AFs), deoxynivalenol (DON), zearalenone (ZEN), fumonisins (FBs) and T-2/HT-2 toxin alter rumen physiology, with special emphasis on dysbiosis, fermentation and epithelial permeability.
1. Anatomy and Histology of the Rumen: Foundations for Understanding Mycotoxin Damage
The rumen occupies most of the left hemibody of the abdominal cavity, with a capacity of 100–250 liters in adult cattle. Its five sacs (dorsal, ventral, cranial, and the caudodorsal and caudoventral blind sacs), delimited by robust muscular pillars as seen in Figure 1, are not mere anatomical divisions: each plays a specific role in the stratification and movement of the contents. The gas dome, the floating fiber layer, the actively fermenting slurry layer, and the ventral liquid layer are organized by density differences and redistributed by the primary and secondary contractions of the reticulo-ruminal cycle (Mattioli, 2003; Yu et al., 2026).
Histologically, the ruminal mucosa is lined by a stratified squamous parakeratinized epithelium, without secretory glands. Its function is exclusively absorptive. The digitiform and linguiform papillae covering the entire internal surface can increase the total absorption area up to 1.0–1.5 m² in adult cattle on a mixed diet, and up to 400% more in animals on highly fermentable diets. Butyrate, one of the three main VFAs, acts as the primary trophic agent on this epithelium: it stimulates proliferation of basal layer cells and maintains papillary architecture. Exposure to mycotoxins that depress butyrate production therefore has direct consequences on the morphological integrity of the absorptive mucosa (de Costa et al., 2007; Vivares et al., 2025).
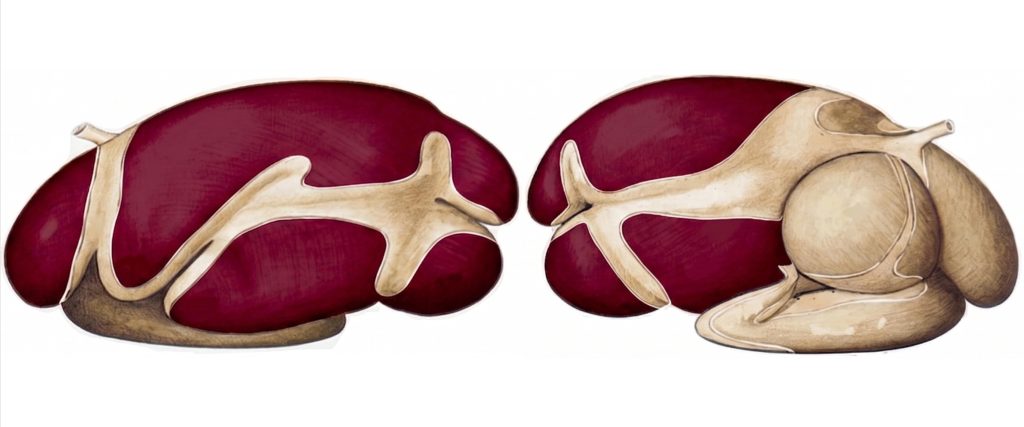
Figure 1. Lateral view of the forestomachs of a ruminant with emphasis of the rumen (colored). Adapted from Popesko (1977).
2. Physiology of Ruminal Fermentation
2.1. The Microbial Ecosystem
Cellulolytic bacteria (Fibrobacter succinogenes, Ruminococcus albus, Ruminococcus flavefaciens) physically attack fiber via cellulosomes, producing endoglucanases and exoglucanases that hydrolyze the β-1,4-glucosidic bonds of cellulose. Anaerobic fungi (Neocallimastix, Piromyces, Orpinomyces) act as pioneers in fiber colonization: their rhizoids physically penetrate the plant cell wall, increasing the surface area for bacterial attack. Ciliated protozoa (Entodinium, Diplodinium) modulate microbial dynamics and act as VFA reservoirs during pH fluctuations.
The final products of fermentation are VFAs (acetate, propionate and butyrate), CO₂ and CH₄. Proportions vary with diet: forages generate 65:20:15 (acetate:propionate:butyrate), while starch-rich concentrates shift fermentation toward propionate (55:30:15). Acetate is the primary substrate for milk fat synthesis; propionate, the main carbon source for hepatic gluconeogenesis; and butyrate, the energy substrate for the ruminal epithelium itself (Annison & Bryden, 1999).
2.2. VFA Absorption and Buffering
The absorption of VFAs occurs mainly by passive diffusion of undissociated forms across the apical membrane of the epithelium. The dissociated fraction is absorbed via SCFA⁻/HCO₃⁻ exchangers, generating a net flow of bicarbonate into the ruminal lumen that buffers pH. Butyrate is metabolized by >90% in the colonocytes themselves, being converted to β-hydroxybutyrate and acetoacetate before being released into the portal circulation. Acetate and propionate flow directly to the liver (Annison & Bryden, 1998).
The rumen buffer system depends critically on saliva: an adult bovine secretes 100–200 liters/day with 120–130 mEq/L of bicarbonate. Rumination, which can occupy 6–10 hours/day, stimulates salivation. Any agent that depresses rumination time directly reduces the buffer supply to the rumen, predisposing to subacute ruminal acidosis (SARA).
3. Impact of Mycotoxins on the Rumen
3.1. Limits of Ruminal Detoxification
The rumen constitutes the first line of defense of the ruminant against ingested mycotoxins, but this capacity for ruminal detoxification has well-defined biological limits. The ruminal microbiota can degrade certain fungal metabolites: ochratoxin A (OTA) is converted to ochratoxin alpha (OTα), of lower toxicity; some trichothecenes such as diacetoxyscirpenol (DAS) are moderately transformed by protozoa. However, aflatoxin B1 (AFB1) and deoxynivalenol (DON) resist the ruminal environment virtually unmodified, maintaining their toxic potential intact (Debevere et al., 2020; Gallo et al., 2024).
Fumonisins (FBs) show minimal ruminal degradation (less than 10%), allowing the full toxin to reach the small intestine without losing its biological activity (Krüger et al., 2022; Gallo et al., 2024). T-2 toxin presents a mixed scenario: although ciliated protozoa can perform moderate metabolism toward less toxic metabolites, this capacity is rapidly saturated at high contamination levels, allowing the intact toxin to pass to the posterior compartments (Debevere et al., 2020).
A particularly concerning case is that of zearalenone (ZEN). The ruminal microbiota converts it mainly to α-zearalenol (α-ZEL), a metabolite with a notably higher estrogenic potency than the original molecule. Instead of detoxifying, the rumen metabolizes the compound, exacerbating reproductive disorders. Studies in Holstein cows fed forage-rich diets showed that ZEN and its metabolites rapidly alter the ruminal microbiome within the first 48 hours of exposure (Hartinger et al., 2023).
3.2. Induced Dysbiosis: Impact on the Microbiome
Mycotoxins that overcome or bypass the ruminal barrier exert direct antimicrobial effects on the resident community. Exposure to FBs and ZEN significantly reduces populations of Lachnospiraceae and Prevotellaceae, two key bacterial families in fiber fermentation and propionate production, respectively. ZEN reduces ruminal pH and the total concentration of short-chain fatty acids (SCFA), compromising the animal’s energy availability. FBs alter the β-diversity of the microbiome and the predicted metagenomic functions of the microbial consortium, documented with only 2 days of exposure (Krüger et al., 2022).
The depression of cellulolytic bacteria by mycotoxins deteriorates fiber degradation and drastically reduces microbial protein synthesis. VFA production falls, generating an energy deficit that the animal cannot compensate for by increasing voluntary intake, which is also depressed. AFB1 reduces gas production, dry matter digestibility and ammonia-nitrogen concentrations in the rumen, evidencing a global deterioration of fermentation (Kemboi et al., 2020). In high-production systems, accelerated passage kinetics reduce ruminal retention time, further limiting detoxification capacity and allowing greater proportions of toxins to pass intact to the other compartments (Gallo et al., 2024).
This microbiological collapse and the resulting pH alteration lead to visible inflammatory lesions in the mucosa, as observed in Figure 2.
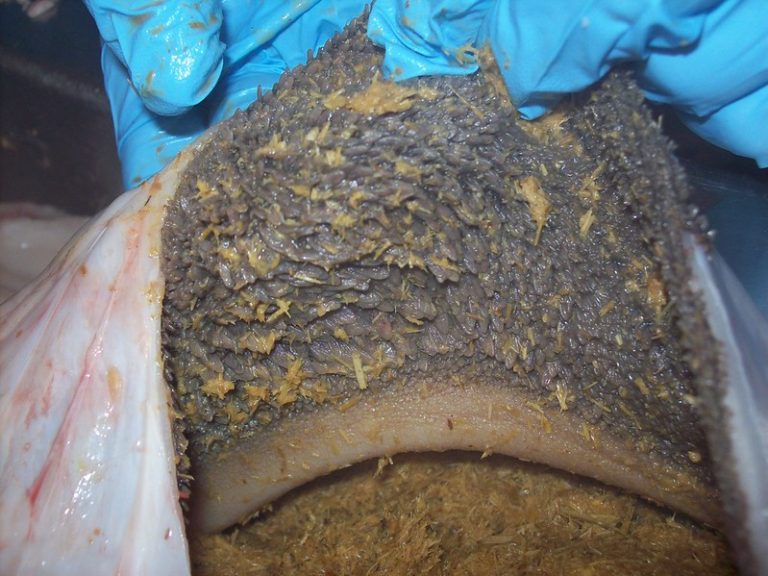
Figure 2. Ruminal mucosa with evident signs of ruminitis. Marked inflammation of the epithelium and alteration of the ruminal papillae can be observed, compromising VFA absorptive capacity.
3.3. Epithelial Permeability and Leaky Gut Syndrome
Mycotoxin-induced dysbiosis also increases ruminal epithelial permeability. The increase in rumen mucosal permeability (Image 1), together with direct damage to the gastrointestinal epithelium, are frequent markers of mycotoxicosis (Antonissen et al., 2014; Gao et al., 2020). Destruction of tight junctions facilitates the translocation of bacterial lipopolysaccharides (LPS) and the toxins themselves into the portal circulation, triggering systemic inflammation and overloading the hepatic detoxification capacity. As detailed in Image 2, this process breaks the natural defensive barrier, allowing the passage of harmful agents via the paracellular pathway.
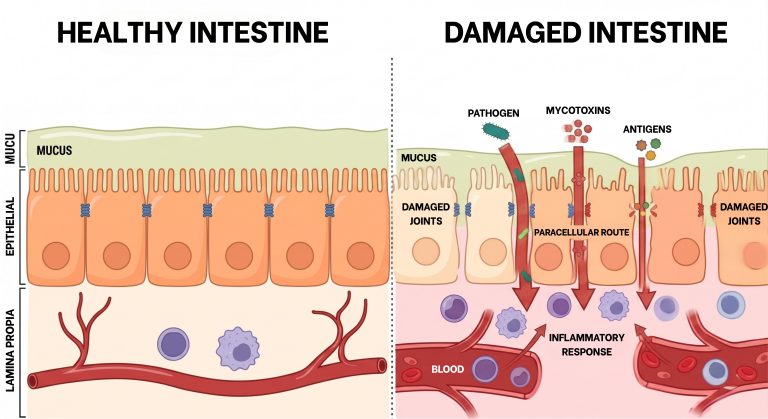
Figure 3. Schematic representation of the loss of tight junction integrity. On the left, a healthy epithelium with a functional barrier; on the right, a damaged epithelium where the paracellular pathway allows translocation of lipopolysaccharides (LPS) and toxins into the bloodstream.
4. Productive and Clinical Consequences
The integration of the above mechanisms produces a metabolic-digestive syndrome that manifests clinically as: reduction in voluntary intake, decline in milk production and daily weight gain, poorer feed conversion efficiency, increased incidence of SARA and its complications (laminitis, hepatitis, hepatic abscesses) and predisposition to infectious diseases due to immunosuppression. In transition cows, the concurrence of negative energy balance and mycotoxigenic dysbiosis exponentially increases the risk of ketosis, retained placenta and metritis (Donkor et al., 2016; Motyka et al., 2025).
Conclusion
The rumen is the first and most important target of mycotoxins in the ruminant. Its microbiome, extraordinarily sensitive to these toxins, can be disrupted even with short exposures and at subchronic concentrations. Ruminal detoxification is real but limited, and in the case of ZEN it can even generate metabolites of greater biological potency. Preventive nutritional management, monitoring of mycotoxins in raw materials, use of advanced bioprotective strategies for mycotoxin mitigation and preservation of microbiome diversity is the strategy most consistent with current scientific knowledge for protecting the health and productivity of the high-producing ruminant.

